UPDATE- Everything in this article has been proven to be most likely correct with Steve Horvath’s new study in mammals that proves aging is programmed and is selected for and conserved by evolution-you can read a fantastic, detailed analysis of his new study here>> https://jefftbowles.com/aging-is-programmed/
Abstract: The last two major areas of evolution that continue to be shrouded in confusion and suffer from unsatisfactory explanation, are sex and aging. A new higher level order of selection is defined as a special form of group selection/species selection termed “predator selection” which allows for a simple solution to the question of the evolutionary purpose of sex and aging. Sex exists to generate and capture beneficial genetic and phenotypic diversity that helps a species survive when confronted with evolving predation. Aging exists to prevent the loss of this beneficial diversity. In the prolonged absence of predation, sex and aging will be lost in a species which will ultimately consist of all clonally reproducing females such as the all-female walking stick insect, whiptail lizards, Brahminy blind snakes, and cave crickets, to name a few. In the absence of predation, programmed aging will be lost and longevity will gradually evolve which can explain all species with exceptionally long lives based on their body size, such as arctic clams 500 years, mouse sized bats 41 years, humans 120 years, box tortoises 130 years, all cave animals, etc. These long-lived examples all have one thing in common: an excellent defense to predation, full body armor, isolation, flight, and extreme intelligence. This article basically summarizes the missing half of Darwin’s theory of evolution.
The last two major areas of evolution that continue to be shrouded in confusion and suffer from unsatisfactory explanation, are sex and aging. In fact, in Dawkins famous book, the Selfish Gene, he throws up his hands and states that the existence of sex is too complicated to explain in his book and “Frankly, I’m going to evade it”. And that was all he could say about sex. Likewise aging got a short mention and a similarly unsatisfactory explanation in his otherwise “comprehensive” book on how the selfish gene drives evolution.

Some theorists try to explain the existence of aging and sex as resulting from group selection where a trait exists in a population that is bad for an individual because it helps the group survive. Most mainstream theorists think conventional group selection is not logical and thus cannot be the force selecting for sex and aging due to the cheater problem and the interbreeding problem. Any interbreeding between groups will quickly destroy differences, and a trait negative to an individual’s fitness should easily be mutated away in a cheater who would quickly spread his or her genes throughout the gene pool. I agree and will not use a conventional group selection argument to demonstrate my case but rather a special form of group selection that is modified to eliminate the cheater/interbreeding problem. So you may by all means read on.
I will make the case here, that the second half of the theory of evolution has yet to be elucidated in any science journal; it will be first done so herein.
I came to the simple solution to the problem of the existence of sex and aging after a 30-year long period of studying aging from all possible angles, publishing several papers about it, and writing a number of books. In the end, it is very simple as you will see.

Most evolutionary theorists tend to take the position that the existence of sex is necessary for most species which somehow helps them avoid extinction. They suggest that most species could not exist without sex. They, for the most part, ignore the cases of sexless animals that reproduce clonally-only such as female walking stick insect (sexless for 100 million years+), female whiptail lizards, some female cave crickets, and the female Brahminy blind snake which exists all over the world and lives underground like a worm for the most part.


Modern day evolutionary theorists generally ignore the very complicated species that can reproduce where the females clone themselves with no need for sex such as the female turkey, the female boa constrictor, the female Komodo dragon, female hammerhead sharks, and many other species of fish. The bottom line is that sex is not required for a species survival yet exists in most animals even though it reduces the ability of the selfish gene to spread by 50% at least! It turns out that after inducing some reverse mutations in mice, scientists were able to create female mice that can reproduce by self-cloning ( 22 April 2004 | Nature | doi:10.1038/news040419-8). It is estimated that it would take about 100-200 mutations to eliminate X-gene imprinting in humans for human females to regain the ability to reproduce clonally! (A Human Virgin Birth Is Technically Possible But Incredibly Unlikely Daniel Engber Pop Sci Nov. 19, 2013)


How about aging? Let’s forget about what is thought of as traditional human aging for now and focus on human menopause, which from an evolutionary perspective is the same thing – something that limits a human’s reproduction (the only human sex that can actually reproduce is the female) and dramatically prevents the spread of her selfish genes. It is obvious to see that menopause is programmed, Dawkins even admits this in his book, so let us ponder it rather than the more gradual aging seen in both sexes of humans.

Before I lose your interest, I will just come out now and tell you how and why sex and aging are selected for by local predation, and then afterwards add some more meat to the bones of this idea. Let us consider a thought experiment to demonstrate the mechanism.
Imagine a local ecosystem with an evolving predator, say a pack of wolves, and a number of species of prey animals that compete with each other to occupy the biomass. Imagine that you have a group of aging and sexually reproducing rabbits that compete with non-aging, asexually-reproducing female ground squirrels to eat the most vegetation and to reproduce the most. Both populations also try to avoid being eaten by the evolving wolves. Now consider that sooner or later a very effective offense evolves amongst the wolves and they can hunt much better than before. Because they can kill and eat one of the identical ground squirrels so easily, they can quickly kill them all – because they are identical!
And thus, the ground squirrel population quickly goes extinct at the local level. However, the aging, sexually reproducing rabbits retain enough genetic and phenotypic diversity that they can evolve a defense to the evolving predation fast enough to avoid local extinction. Aging, say menopause, prevents any one female rabbit from reproducing too much and homogenizing the gene pool with too many of the same genes. AGING RETAINS GENE POOL DIVERSITY. Sexual reproduction, on the other hand, creates phenotypic diversity and allows the various mutations that occur in different individuals to come together in a single individual much more quickly as compared to clonally-reproduced individuals where a large number of beneficial mutations would have to occur simultaneously (much more unlikely) in an individual to create a similar beneficial phenotype. SEX CREATES PHENOTYPIC DIVERSITY. And as we will soon see the male sex type also CREATES GENE POOL DIVERSITY.

Vs

Vs.

So the bottom line is that the aging and sexually reproducing rabbits will be much more likely to survive local (or novel) evolving predation and in many cases will eventually migrate to other ecosystems from time to time and will be able to recolonize areas that were once occupied by non-aging asexual rabbits that have become extinct due to encountering evolving predation in their respective ecosystems. (The sexual, aging rabbits can also migrate and replace non-aging asexually reproducing ground squirrels or any other non-aging or asexual animal that might compete with the sexual, aging rabbits for habitat). Also, predators can migrate to new ecosystems and wreak havoc and we see that they can and do hunt various species to extinction if the indigenous prey species cannot evolve new defenses quickly enough (the problem of invasive species).
In fact, if you consider that within an ecosystem most animals and organisms can kill or be killed by many of the species in the ecosystem, you could say that aging and sex evolve as emergent properties of local ecosytem dynamics from the interconnected web of groups competing for eating and not being eaten. The elephants and lions evolve aging and sex for the same reason that ants and plants do.
So in the end, the selfish gene theory is actually correct, and the entire world would consist of all asexually reproducing non-aging female clones if not for evolving predation at the local ecosystem level. The danger of extinction caused by losing sex or aging is so great that evolution has somehow gone to great lengths to prevent it from happening, including the creation of multiple, redundant aging systems and multiple mutations that prevent the reacquisition of the ability for females to reproduce asexually. Thus, it is very rare to see asexually reproducing or non-aging species – but they do occur from time to time, and are at great risk of extinction if a predator evolves a new way to kill them.
Does this sound like conventional group selection? Maybe a little, but in fact it is a level of selection one order higher than group selection; it is actually a form of species selection at the local level, that occurs everywhere all the time for the most part and is the reason why sex and aging are almost ubiquitous on the planet. You could call it modified group selection if you wanted to as it still depends on competition between groups of different species at the local level.
This new form of species selection is quite different from the idea of species selection as put forth by Stephen Jay Gould (and others like Williams’ version called clade selection) where he describes how some species survive major planetary catastrophes while other species don’t. The form of species (or modified group) selection I am describing happens on a small scale at the ecosystem level, and all the time; while Gould’s idea of species selection happens on a planetary scale and maybe once every 50 million years or so. Very different from my idea of species selection – so different that I would like to coin the term predator selection to describe this special form of species selection.

Not convinced? Let’s put some more meat on the bones. How about this. Males, and male sex traits exist solely as a defense to evolving predation and will disappear eventually if a species is no longer preyed upon significantly for prolonged periods. This is just as it should be based on the selfish gene theory as there is a huge cost in evolutionary fitness for having 50% of the population being male. Males cannot reproduce and cause the diminishment of 50% of the spread of the female’s genes. A perfect situation for the “cheater” theory to come into play, a female that re-evolves the ability to reproduce clonally to quickly spread 100% of her selfish genes as opposed to just 50% will have a huge advantage and over time cannot be resisted.
And what is the primary purpose of male sex traits such as bright colors, loud songs, crazy dances?
Not to attract females. That purpose is only secondary.
The primary purpose of most male sex traits are to attract predators so that only the most fit in the presence of predation will survive the burden of living with loud attractive targets on their backs.
And that male with the winning gene combination who survives, gets to impregnate all the camouflaged females he can find. (It’s good to be the king!)

Also, there are some other male sex traits that are a little less obvious, but are attractive to females, and serve the same purpose such as body size, antler length, feather length, even gray hair and beard length! These indicate a male’s age, and thus the number of predator encounters he has likely survived. Females are impressed by age markers as well as predator attractants.

How about aging? Most animals age and die at a rate related to their body size, a mouse – 2 years, a dog – 15 years, a horse – 40 years etc., but there are a number of animals that live much longer than expected based on their body size such as a mouse-sized bat named myotis – 41 years, an arctic (small) clam 500 years, cave salamander 170 years, humans 120 years, box tortoises 130 years, deep dwelling fish 125 years, lobsters 220 years, and some birds like parrots 90 years!
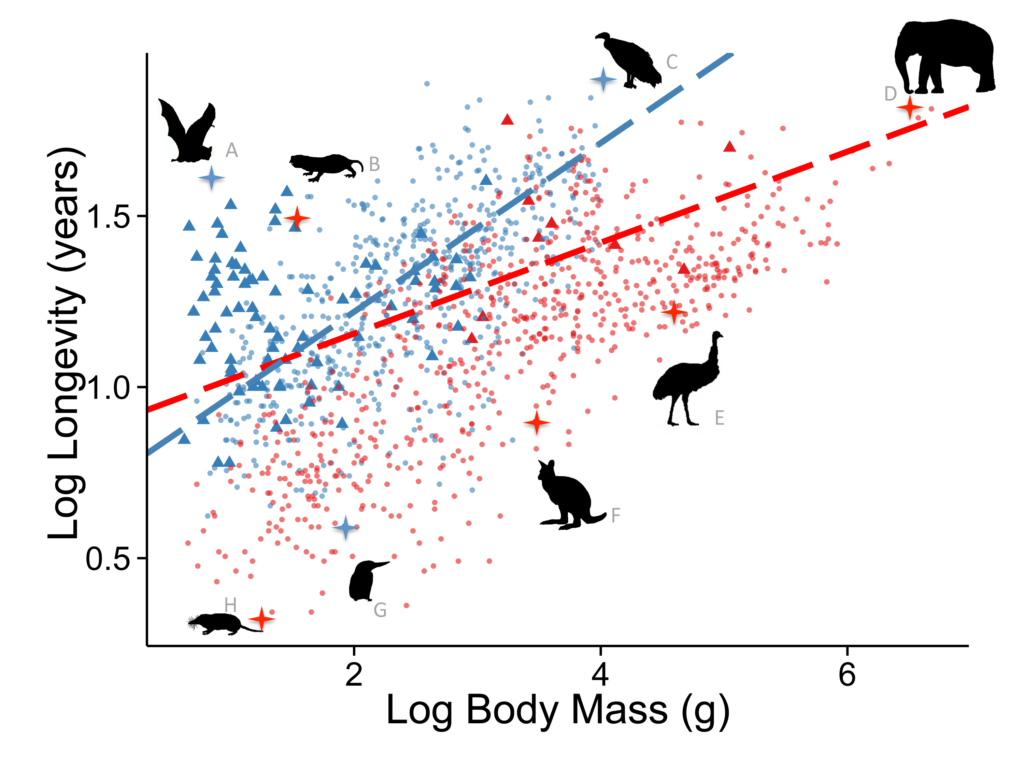
What do all these long living animals have in common? A great defense to predation, whether it is full body armor, isolation, flight, or extreme intelligence. Once an effective defense to predation evolves then aging can be selected against but it is lost gradually from an evolutionary point of view due to the redundant and multiple aging systems embedded in their genomes (as described in some of my previously published journal articles).
The only remaining question I still had after writing this article is why are there male sex types in plants? What good is a plant that cannot reproduce? And how can a male plant evolve a better way to defend against predators when he can’t move? And after a little thought I realized there is another purpose to the male sex other than attracting predators.
Another important purpose of male of all species, whether plant or animal, is to generate and be the bearer of a much larger share of genetic germline mutations (probably by the actions of the intronless SRY gene) , that are usually detrimental but occasionally beneficial to surviving evolving predation. The male is basically the testing ground for new germline mutations in the face of predation. In fact, if you do a little research on the subject you will discover the existence of male mutation bias that occurs in almost all species including plants. In humans, the male germline mutation rate is 5X higher than the mutation rate in females!
Let us summarize with a few new observations about evolution:
SEX EXISTS TO RAPIDLY CREATE BENEFICAL DIVERSITY IN GENES AND PHENOTYPES–
Sex exists to rapidly increase beneficial phenotypic diversity in the face of evolving local predation-via recombination.
The male sex exists to generate a high number of germline mutations that are usually detrimental, but occasionally beneficial with respect to surviving evolving predation – via the male mutation bias.
Most male sex traits exist to attract predators who weed out the detrimental mutations and phenotypes, and select for the smaller number of beneficial mutations. Male traits are attractive to females but also attractive to predators – sexual selection.
Some male sex traits exist to inform females that their male genes are older, predator-tested and superior to younger individuals’ genes (age markers).
AGING EXISTS TO PREVENT THE LOSS OF BENEFICAL DIVERSITY IN GENES AND PHENOTYPES–
Aging exists to prevent the loss of beneficial genetic diversity by restricting the amount of any one individual’s contribution to the gene pool.
And finally another interesting insight- if we assume the phenomenon of menopause evolved to stop/limit a female’s contribution to the gene pool to preserve the gene pool’s diversity, let us look at some other parameters that are linked to menopause (aka declining litter size in other animals) :
These folowing factors are all linked across species…Length of pregnancy (gestation time), age at menarche (puberty), litter size, age of decline in litter size, and maximum lifespan. When you increase the maximum life span in a species, it pulls all the other parameters along with it: It increases the gestation period (this decreases the number of offspring possible in a lifetime) it increases the age of menarche (puberty) (this also decreases the number of offspring possible in a lifetime), it decreases litter size (this also decreases the number of offspring possible in a lifetime)- starting to see a pattern here? All these things reduce total possible life time reproduction potential as lifespan increases.
And finally we can start to make sense of homosexuality which no biologist to my knowledge has ever tried to explain in evolutionary terms even though it occurs throughout the animal kingdom from humans all the way down to insects! If you do a little research you will find that homosexual offspring are born to mothers who were stressed during pregnancy at a certain time of brain development in the fetus. Stress in the evolutionary past likely came from encounters with predators that were survived by the mother. The stress caused the elevation of the stress homrone cortisol which impacted the development of the embryo by suppressing the sex steroids that alter brain development. So how could this be selected for at the species level?
Answer: Homosexuality basically acts as birth control for evolutionarily “unfit” mothers. While a female mammal nurses her young she is no longer able to get pregnant. She then will spend time and resources raising offspring that will likely never reproduce, thus preventing her unfit genes, at least temproarily, from being added to the gene pool. In humans, various forms of “fake” stress , like smoking, drug abuse, and/or drinking can lead to elevated cortisol levels and thus to homosexual offspring. Quitting smoking is very hard for some people. It would not be too much of a stretch to imagine that some pregnant human females hide their smoking from others while pregnant. I write in much more detail about this in my book What Darwin Could Not See- The Missing Half of the Theory.
This is the first journal article I have written since 2000 and I expect it to be the last. If you are interested on more detailed versions of these arguments, and further extension of this viewpoint to explain additional things like the evolutionary purpose of homosexuality, delayed fertility in longer living species, sex changing animals, and the biochemistry of aging, please see my books available at amazon.com (This was submitted to and rejected by The Journal of Theoretical Biology)
Jeff T Bowles
Glad you liked it! PLease share it with your friends.
318309 193240Thank you, Ive recently been searching for info about this subject for ages and yours will be the greatest Ive discovered out so far.
Jeff Bowles is a brilliant outside the box scientist. I consider him among the great thinkers on evolutionary biology, aging and why certain hormones are protective against the oxidative stressors of aging. He breaks through presumptions.
Phyllis J. Bronson, Ph.D.
Phyllisbronsonphd.com