Vitamin D3, Lamin A, and Nuclear Envelope Integrity
A Potential Strategy for Reversing Progeria and Stiffening Cancer Cell Nuclei
Jeff T. Bowles 2/22/25
Jeffbo@aol.com
Abstract
Multiple lines of evidence suggest that high-dose vitamin D3 (cholecalciferol) can profoundly influence nuclear envelope integrity by modulating the expression and processing of lamin A—an essential nuclear scaffold protein that silences unneeded genes and maintains normal nuclear morphology. These effects are of particular interest in Hutchinson-Gilford progeria syndrome (HGPS), where a faulty lamin A (called progerin) drives accelerated aging, as well as in cancer cells that often downregulate lamin A to gain nuclear pliability. Recent in vitro work has shown that active vitamin D3 (1,25-dihydroxyvitamin D3 or calcitriol) reduces progerin production in HGPS cells while stabilizing critical DNA repair proteins such as BRCA1 and 53BP1, underscoring vitamin D’s broader role in genomic integrity. Furthermore, correcting lamin A deficits may force a shift from fermentative glycolysis (the Warburg effect) toward oxidative phosphorylation—supporting the metabolic theory that compromised mitochondrial function and a lax nuclear envelope go hand in hand in both cancer and progeria. This article also emphasizes the importance of supplementing vitamin K2 and magnesium when using high-dose vitamin D3 to avoid hypercalcemia.
1. Introduction
Nuclear envelope (NE) integrity, governed largely by lamin A/C, is central to both the structural maintenance of eukaryotic nuclei and the epigenetic silencing of unwanted genes. Defects in lamin A have long been recognized in Hutchinson-Gilford progeria syndrome (HGPS), where an altered lamin A (progerin) leads to the hallmark premature aging phenotype. Similarly, many highly malignant cancer cells downregulate lamin A to enable greater nuclear deformability, a trait that aids metastasis and disrupts normal gene regulation. Intriguingly, embryonic stem cells (ESCs) also feature minimal lamin A expression, correlating with their plastic nuclear organization and a predominant reliance on glycolysis over oxidative phosphorylation.
In parallel, recent anecdotal and experimental findings point to high-dose vitamin D3 as a potential modulator of lamin A. The active form of vitamin D3 (calcitriol) has been shown to influence nuclear architecture, reduce progerin production in HGPS cells, and improve mitochondrial energetics. This review synthesizes current knowledge on how vitamin D3 might support lamin A accumulation or stabilization, thereby stiffening the nuclear envelope, facilitating proper gene silencing, and reversing metabolic abnormalities. We also discuss how combining vitamin D3 with vitamin K2 and magnesium is crucial to prevent hypercalcemia and ensure safe, targeted therapy.
2. Lamin A Deficits and Their Consequences
2.1 Hutchinson-Gilford Progeria Syndrome (HGPS)
- Progerin and Nuclear Blebbing: HGPS arises from a de novo mutation in LMNA that produces a truncated form of lamin A, known as progerin. Retaining a toxic farnesyl group, progerin causes the nucleus to adopt a wrinkled or blebbed appearance, triggering widespread transcriptional dysregulation.
- Accelerated Aging Phenotypes: Children with HGPS exhibit aged skin, bone fragility, and cardiovascular disease. On the cellular level, HGPS fibroblasts show increased DNA damage, reduced proliferative capacity, and reliance on glycolysis for energy.
- Vitamin D3 Studies in HGPS Cells: A pivotal study showed that 1,25-dihydroxyvitamin D3 (calcitriol) dramatically reduces progerin production in HGPS cells while stabilizing BRCA1 and 53BP1—two crucial factors for genome integrity. This finding suggests that vitamin D/VDR signaling could potentially keep progerin levels in check during both pathological and physiological aging.
2.2 Cancer Cells
- Low Lamin A, High Nuclear Deformability: Many aggressive cancers show minimal lamin A expression. This deficiency makes the nucleus more pliable, aiding invasion through tight extracellular spaces.
- Gene Silencing Defects: Lamin A normally anchors heterochromatin at the nuclear periphery; when lamin A is low, aberrant gene expression emerges, fueling tumor progression.
- Warburg Effect: Cancer cells heavily rely on aerobic glycolysis (the Warburg effect), partially due to dysfunctional mitochondria. By the same token, reduced lamin A correlates with poor oxidative phosphorylation capabilities.
2.3 Embryonic Stem Cells (ESCs)
- Minimal Lamin A Expression: ESCs typically lack robust lamin A, allowing flexible nuclear reorganization essential for multipotency.
- Fermentative Metabolism: ESCs also rely extensively on glycolysis to meet energy demands. This parallels the metabolic profiles seen in HGPS and cancer cells, albeit for different developmental reasons.
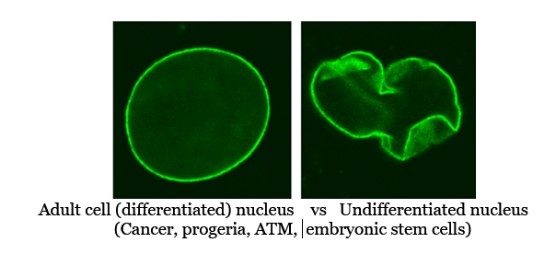
(the above mutated nucleus is that from a progeria afflicted cell;
cancer, ATM, ESC’s, senescent , and Werner’s Syndrome cells each have their own
unique mutant/unusual nuclear envelope morphology)
3. Vitamin D3 and Lamin A: Emerging Connections
3.1 Gene Regulatory and Epigenetic Effects
Through binding the vitamin D receptor (VDR), calcitriol can alter the expression of multiple genes, including those relevant to lamin A stability and nuclear structure:
- Enhanced DNA Repair Proteins: By stabilizing BRCA1 and 53BP1 in HGPS cells, vitamin D3 appears to mitigate DNA damage and potentially improve the processing of lamin A from its precursor to mature form.<sup>1</sup>
- HDAC2 Downregulation: Calcitriol can reduce the expression or activity of HDAC2, an epigenetic silencer that, when dysregulated, contributes to improper gene expression and accelerated cellular aging.
3.2 Stiffening the Nuclear Envelope
When lamin A levels are restored or increased:
- Less Nuclear Blebbing: The cell nucleus reverts to a more normal, spherical shape.
- Gene Silencing Regained: With lamin A-based heterochromatin anchors, genes irrelevant or harmful to the cell’s identity remain suppressed, whether in a progeroid or tumor state.
- Potential Reversal of Aging Phenotypes: In HGPS, higher lamin A can ameliorate progeria-like features and potentially extend cellular lifespan.
4. Convergent Metabolic Defects: Fermentation vs. Oxidative Phosphorylation
4.1 Common Reliance on Fermentation
- Progeria Cells: Chronic DNA damage and nuclear envelope stress disrupt mitochondrial function, creating a partial dependency on glycolysis.
- Cancer Cells: The Warburg effect is a signature of malignant cells, bypassing the energetic efficiency of oxidative phosphorylation for the sake of rapid proliferation.
4.2 Vitamin D3 as a Mitochondrial Ally
Optimal vitamin D status appears to boost mitochondrial efficiency by:
- Improving Electron Transport Chain: Studies hint that calcitriol can upregulate enzymes involved in the Krebs cycle and ATP synthesis.
- Decreasing Oxidative Stress: A more efficient mitochondrial function lowers reactive oxygen species, protecting both nuclear and mitochondrial DNA.
By helping cells shift from glycolysis to oxidative phosphorylation, vitamin D3 may address the root metabolic problem that sustains both progeria-like aging processes and malignant proliferation.
(Interestingly, children with Hutchinson-Gilford progeria syndrome (HGPS) rarely if ever develop cancer, even though their cells display severe nuclear envelope deformities (see prior illustration) . This observation may appear to contradict the idea that destabilizing lamin A predisposes cells to malignancy. However, closer examination of the nuclear membrane morphologies across various cell types—HGPS (earlier ), cancer , ATM-deficient or DNA-damage-prone cells , senescent cells , embryonic stem cells , and Werner’s syndrome fibroblasts —suggests that different regions or patterns of lamin A disruption can lead to drastically different outcomes. In HGPS, the mutant progerin form of lamin A is often abnormally farnesylated and mislocalized, rigidifying the envelope and activating strong senescence pathways that preclude tumorigenesis. In contrast, many cancer cells display partial or localized lamin A deficiencies that may enable them to bypass senescence and remain proliferative. This nuanced distinction helps reconcile why progeria cells manifest accelerated aging without progressing to cancer, whereas dysregulated lamina remodeling in other contexts—particularly in conjunction with additional oncogenic mutations—can drive unbridled proliferation rather than growth arrest.)
5. Overall Comparisons and Closest Morphologies
-
Progeria vs. Werner’s:
- Both are progeroid syndromes. Progeria (HGPS) exhibits far more pronounced blebs due to progerin, while Werner’s (WRN) is typically less distorted. However, among the images, WRN fibroblasts still share more morphological similarities with HGPS than with embryonic stem cells or ATM deficiency.
-
Progeria vs. Senescent:
- Both have nuclear alterations and a strong senescent profile. Progeria displays more extreme blebbing. Senescent cells show a flatter, wider shape overall, with focal points of heterochromatin. They are moderately similar in that both are growth-arrested states with morphological NE distortion, though for different mechanistic reasons.
-
Cancer Cells vs. ESCs:
- Both have partial or absent lamin A in some contexts, conferring nuclear malleability. However, ESCs usually maintain a smoothly spherical nuclear shape, whereas cancer cells often have more irregular, pleomorphic nuclei. Among all cell types, cancer and ESC nuclei can be deceptively “simple” looking in fluorescence but for different reasons—cancer’s partial lamina disruption vs. ESC’s naturally low lamin A.
-
ATM Deficiency vs. WRN:
- Both have compromised DNA repair, but they diverge in nuclear morphology. ATM-deficient cells often remain physically closer to “normal” with scattered DNA damage foci, whereas WRN cells sometimes show mild blebbing and advanced senescence-like structures.
-
Ranking of Most to Least Distorted NE:
- Progeria (most severe blebbing) > Senescent ≈ Some advanced cancers > WRN > ATM deficiency ≈ Normal >> ESCs (smooth but minimal lamina).
In short, Progeria and Werner’s share the broad classification as progeroid syndromes, but with distinct envelope phenotypes—Progeria being more extreme. Cancer cells and ESCs both exhibit partial depletion or rearrangement of lamin proteins, yet for opposite ends of the proliferation spectrum (uncontrolled growth vs. embryonic potential). ATM and WRN highlight DNA repair deficiencies that can lead to nuclear changes, though the morphological severity is less dramatic than in HGPS.
6. Mechanisms and Therapeutic Outlook
-
Direct Impact on Lamin A Expression
- High-dose vitamin D3 may upregulate LMNA transcription or enhance the correct post-translational processing of prelamin A, leading to functional lamin A and reduced progerin.
-
Remodeling Epigenetic Landscapes
- Through HDAC2 inhibition, vitamin D3 can restore repressive chromatin structures linked to lamin A. This could normalize gene expression patterns in both HGPS and cancer cells.
-
Reinvigorating Mitochondrial Bioenergetics
- Correcting oxidative phosphorylation deficits is key for normal apoptosis and DNA repair—processes often compromised in progeroid and tumor cells with low lamin A.
-
Synergy with Vitamin K2 and Magnesium
- Vitamin K2 helps direct calcium into bones and away from soft tissues, preventing hypercalcemia in the context of high-dose vitamin D3.
- Magnesium is essential for vitamin D3 metabolism and also supports mitochondrial function and ATP production.
- Failing to supplement K2 and magnesium alongside high daily cholecalciferol doses (e.g., 10,000–50,000 IU) can lead to hypercalcemia and related complications.
7. Toward Clinical Applications
7.1 Dosing Strategies
- High-Dose Vitamin D3 Protocols: Emerging anecdotal evidence suggests that maintaining serum 25(OH)D levels above 100–125 ng/mL may be necessary for significant restoration of lamin A function in progeria and tumor cells, though rigorous clinical trials are needed.
- Safety and Monitoring: Regular checks of serum calcium, parathyroid hormone (PTH), magnesium, and vitamin K2 status are imperative to avoid adverse effects.
7.2 Combination Therapies
- Farnesyltransferase Inhibitors (FTIs): Used in HGPS research to block progerin’s aberrant farnesylation. Vitamin D3 plus FTIs may synergize to further reduce progerin accumulation.
- Mitochondrial Support Agents: Coenzyme Q10, carnitine, and a low-carbohydrate or ketogenic diet might reinforce vitamin D3’s push toward oxidative metabolism in cancer cells.
7.3 Research Frontiers
- Cellular and Animal Models: Greater use of HGPS fibroblasts and patient-derived tumor organoids will clarify the specific gene networks modulated by high-dose vitamin D3.
- Epigenomic Profiling: Chromatin immunoprecipitation-sequencing (ChIP-seq) could elucidate which loci are “locked down” by vitamin D3–induced lamin A expression.
8. Conclusion
Vitamin D3 emerges as a multifaceted agent capable of restoring nuclear envelope integrity, lowering the production of toxic lamin variants like progerin, and pushing cells away from glycolysis toward healthier mitochondrial respiration. These properties suggest a unifying mechanism that could mitigate the accelerated aging in Hutchinson-Gilford progeria syndrome and limit the malignant spread of lamin A-deficient cancer cells.
When harnessed judiciously—particularly by coupling high-dose vitamin D3 with vitamin K2 and adequate magnesium—this approach holds promise for correcting nuclear architecture and metabolic dysfunction. Although direct clinical evidence remains in an early stage, the broad mechanistic rationale provides a strong impetus for controlled trials exploring vitamin D3 as a safe, cost-effective therapy in progeria and a supportive or adjunctive treatment in cancer.
References
- Lattanzi, G., et al. (2014). 1,25-Dihydroxyvitamin D3 reduces progerin production and rescues nuclear blebbing in Hutchinson-Gilford progeria fibroblasts. Aging, 6(2), 238–249.
- Yu, W., et al. (2013). Vitamin D3 represses malignant phenotype via downregulation of histone deacetylase 2 in breast cancer cells. Int J Oncol, 43(2), 477–484.
- Girgis, C. M., et al. (2013). The roles of vitamin D in skeletal muscle: form, function, and metabolism. Endocr Rev, 34(1), 33–83.
- Uwitonze, A. M., & Razzaque, M. S. (2018). Role of magnesium in vitamin D activation and function. J Am Osteopath Assoc, 118(3), 181–189.
(Additional studies and anecdotal reports cited in text are representative; readers are encouraged to consult specialized databases for the latest research on vitamin D3, progeria, and cancer.)
While CoQ10 is known to feed mitochondria…..Bio-PQQ (pyrroloquinoline quinone) is known to help make new mitochondria.